This thread is not about mathematics. It's about Microtubules..")
But it is . Your Philosophy about Mathematics being the essence of the Universe , affects, effects , causes , Everything . Is Involved in Everything . Including Microtubules .

This thread is not about mathematics. It's about Microtubules..
https://www.sciencedirect.com/science/article/pii/S0161589019300951#Microtubules (MTs) are tubular polymers of tubulin that are highly dynamic and found throughout the cytoplasm and play a role in the innate and adaptive immune systems.
Studies suggest an essential role for MTs in the gut.
Here, we propose a model that represents gut MTs as potential targets for immunotherapy.
OK, microtubules are nano-scale computers that process electro-chemical information (qubits)But it is . Your Philosophy about Mathematics being the essence of the Universe , affects, effects , causes , Everything . Is Involved in Everything . Including Microtubules .
The scientific approach to understand the nature of consciousness revolves around the study of human brain. Neurobiological studies that compare the nervous system of different species have accorded highest place to the humans on account of various factors that include a highly developed cortical area comprising of approximately 100 billion neurons, that are intrinsically connected to form a highly complex network. Quantum theories of consciousness are based on mathematical abstraction and Penrose-Hameroff Orch-OR Theory is one of the most promising ones.
https://arxiv.org/ftp/arxiv/papers/1505/1505.00774.pdf#Inspired by Penrose-Hameroff Orch-OR Theory, Behrman et. al. (Behrman, 2006) have simulated a quantum Hopfield neural network with the structure of a microtubule. They have used an extremely simplified model of the tubulin dimers with each dimer represented simply as a qubit, a single quantum two-state system. The extension of this model to n-dimensional quantum states, or n-qudits presented in this work holds considerable promise for even higher mathematical abstraction in modelling consciousness systems.
Microtubules (MTs) are long cylindrical structures of the cytoskeleton that control cell division, intracellular transport, and the shape of cells. MTs also form bundles, which are particularly prominent in neurons, where they help define axons and dendrites. MTs are bio-electrochemical transistors that form nonlinear electrical transmission lines. However, the electrical properties of most MT structures remain largely unknown. Here we show that bundles of brain MTs spontaneously generate electrical oscillations and bursts of electrical activity similar to action potentials.
Under intracellular-like conditions, voltage-clamped MT bundles displayed electrical oscillations with a prominent fundamental frequency at 39 Hz that progressed through various periodic regimes. The electrical oscillations represented, in average, a 258% change in the ionic conductance of the MT structures. Interestingly, voltage-clamped membrane-permeabilized neurites of cultured mouse hippocampal neurons were also capable of both, generating electrical oscillations, and conducting the electrical signals along the length of the structure.
https://www.nature.com/articles/s41598-018-30453-2Our findings indicate that electrical oscillations are an intrinsic property of brain MT bundles, which may have important implications in the control of various neuronal functions, including the gating and regulation of cytoskeleton-regulated excitable ion channels and electrical activity that may aid and extend to higher brain functions such as memory and consciousness.

Mirror neurons are neurons in your brain that fire when you see something happen to someone or when someone performs an action. Mirror neurons are why you flinch when you unexpectedly see someone getting punched, or why your hands may twitch if you see someone grab an object. These neurons make you experience the same sensations as the person who performed the action in the first place, so it’s as if you were punched in the face or grabbed the object.
Mirror neurons were actually discovered accidentally. In the 1990s, Italian researchers hooked macaques up to electrodes so that their brain activity could be monitored. One day, when a researcher reached out to grab some food during his lunch break, he noticed that one of the monkeys was watching him and neurons were activated in his prefrontal cortex, the area of the brain that controls movement. The brain activity was exactly as if the macaque had grabbed the food himself.
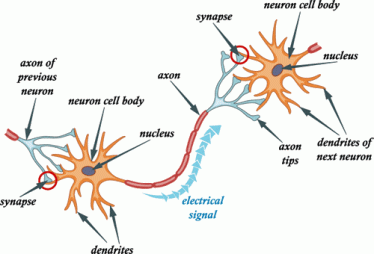
https://issciencejournalblog.wordpress.com/2017/03/15/mirror-neurons-the-gateway-to-empathy/This led to further experimentation in 1995, where a study found that when people watched someone grabbing an object, their hand muscles instinctively tensed as if in preparation for grabbing that object themselves, which proved the existence of mirror neurons.
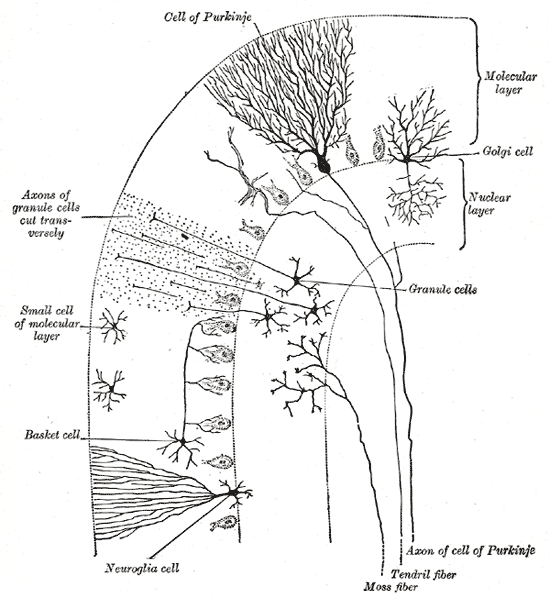
In humans, Purkinje cells can be harmed by a variety causes: toxic exposure, e.g. to alcohol or lithium; autoimmune diseases; genetic mutations causing spinocerebellar ataxias, gluten ataxia, Unverricht-Lundborg disease, or autism; and neurodegenerative diseases that are not known to have a genetic basis, such as the cerebellar type of multiple system atrophy or sporadic ataxias.[34][35]
https://en.wikipedia.org/wiki/Purkinje_cellGluten ataxia is an autoimmune disease triggered by the ingestion of gluten.[36] The death of Purkinje cells as a result of gluten exposure is irreversible. Early diagnosis and treatment with a gluten-free diet can improve ataxia and prevent its progression.[34][37] Less than 10% of people with gluten ataxia present any gastrointestinal symptom, yet about 40% have intestinal damage.[37] It accounts for 40% of ataxias of unknown origin and 15% of all ataxias.[37]
Simple. Because they're not at the heart of all neural cells, nor are they responsible for dendritic growth.But nowhere in this in-depth analysis of Purkinje cells is there any mention of the "microtubules" (except for a passing mention of mitochondria), which are at the heart of all neural cells and responsible for cellular (dendritic) growth and shrinkage, as in Alzheimers disease.
Yep. I remember back when people thought miasmic humours were the cause of all disease. It was a neat theory that tied everything together simply without requiring a laundry list of viruses, bacteria, prions, parasites and amoebas. It bound everything together! Nowadays doctors compartmentalize and treat all those things as separate phenomena with separate pathogens - which is why we've cured many of them.This is why I try to find "common denominators" that bind ecerything together in reality rather than the compartmentalization that treats everything as separate phenomena......
We disagree. Microtubules provide the mitotic spindle for all cell division (especially neural cells) in the first place .Simple. Because they're not at the heart of all neural cells, nor are they responsible for dendritic growth.
Even earlier diseases were attributed to demons...?Yep. I remember back when people thought miasmic humours were the cause of all disease. It was a neat theory that tied everything together simply without requiring a laundry list of viruses, bacteria, prions, parasites and amoebas. It bound everything together!
Of course, but we are not talking about treating specific diseases and their unique demands. Treat the comment in context of the subject.Nowadays doctors compartmentalize and treat all those things as separate phenomena with separate pathogens - which is why we've cured many of them.
 Micaela Lasser,
Micaela Lasser, 
 Laura Anne Lowery*
Laura Anne Lowery*Neurons depend on the highly dynamic microtubule (MT) cytoskeleton for many different processes during early embryonic development including cell division and migration, intracellular trafficking and signal transduction, as well as proper axon guidance and synapse formation.
The coordination and support from MTs is crucial for newly formed neurons to migrate appropriately in order to establish neural connections. Once connections are made, MTs provide structural integrity and support to maintain neural connectivity throughout development. Abnormalities in neural migration and connectivity due to genetic mutations of MT-associated proteins can lead to detrimental developmental defects.
Growing evidence suggests that these mutations are associated with many different neurodevelopmental disorders, including intellectual disabilities (ID) and autism spectrum disorders (ASD). In this review article, we highlight the crucial role of the MT cytoskeleton in the context of neurodevelopment and summarize genetic mutations of various MT related proteins that may underlie or contribute to neurodevelopmental disorders. Menon and Gupton, 2016; Pacheco and Gallo, 2016; Kirkcaldie and Dwyer, 2017).
Newly formed neurons face many challenges as they undergo dramatic changes in shape and migrate their way through the extracellular terrain in order to establish connections with other cells. Specifically, dynamic MTs play pivotal roles in creating cell polarity, as well as aiding in neural migration in order to establish appropriate neural connectivity throughout development and into adulthood.
The elaborate MT network is integral to facilitate numerous morphological and functional processes during neurodevelopment, including cell proliferation, differentiation and migration, as well as accurate axon guidance and dendrite arborization. The organization and remodeling of the MT network is also essential for developing neurons to form axons, dendrites and assemble synapses.
Moreover, in mature neurons, MTs continue to maintain the structure of axons and dendrites, and serve as tracks for intracellular trafficking, allowing motor proteins to deliver specific cargoes within the cell.
As brain development relies heavily on proper MT function, defects in the MT cytoskeleton can lead to detrimental effects on neural proliferation, migration and connectivity. Over the last several years, numerous studies have identified mutations within genes coding for proteins that interact with and directly modulate the structure and function of the MT cytoskeleton.
Many of these MT-associated mutations have been linked to various neurodevelopmental disorders including lissencephaly, polymicrogyria, autism spectrum disorders (ASD) and intellectual disabilities (ID; Srivastava and Schwartz, 2014; Chakraborti et al., 2016; Stouffer et al., 2016). The regulation of the MT cytoskeleton during specific stages of brain development still remains an active topic of research. In this review article, we highlight various studies that illustrate important functions of the MT cytoskeleton that contribute to proper neural development and how genetic mutations within MT-related proteins can alter these crucial functions that may lead to disorders of neural development. 1; Akhmanova and Steinmetz, 2008).
MTs are extremely dynamic structures, existing in either a growing state (polymerization) or shrinking state (depolymerization). The plus ends of MTs can rapidly switch between these two states, going from growth to shrinkage (catastrophe), or from shrinkage to growth (rescue), a process called “dynamic instability” (Mitchison and Kirschner, 1984). Developing neurons depend on this stochastically dynamic nature of the MT cytoskeleton in order to remodel their shape, proliferate and migrate, as well as other processes during different phases of neural development, as described in more detail below.
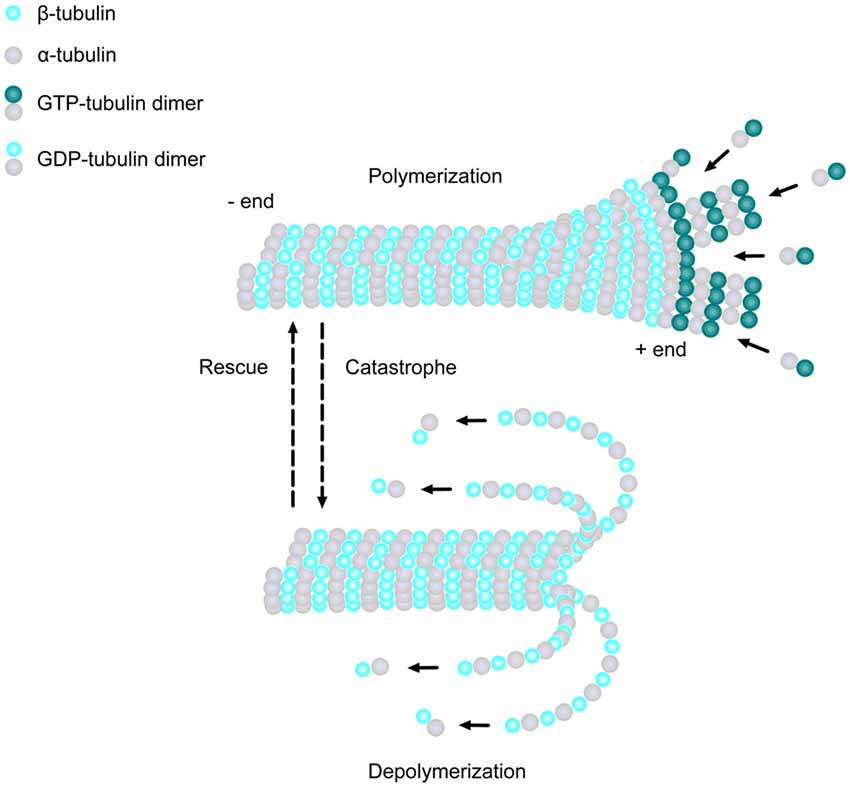
Neurons begin their development as spherical, unpolarized cells, with MTs emanating from MT organizing centers (MTOCs), such as the centrosome, and are nucleated by the γ-tubulin ring complex (γ-TuRC; Kuijpers and Hoogenraad, 2011). This structure acts as a template for the α- and β-tubulin dimers to begin polymerization and is the cap of the minus end as the MT plus end grows away from the MTOC (Kuijpers and Hoogenraad, 2011). Once differentiation occurs, neurons form multiple processes, termed neurites, which extend from the spherical cell body and elongate to form thin protrusions (Götz and Huttner, 2005). MTs are one of the major players which influence the formation of neurites by creating small bundles that invade lamellipodia in multiple directions (Götz and Huttner, 2005).
It has been suggested that MT sliding may play a key role in initiating neurite formation. One study demonstrated that MT-associated protein 2c (MAP2c) could stabilize MT bundles in vitro, which then rapidly move toward the cell periphery, prompting protrusions via a dynein-driven force (Dehmelt et al., 2006). It has also been shown that neurite formation can be induced by the MT motor protein, kinesin-1, which drives MT sliding and generates a mechanical force on the cell membrane (Lu et al., 2013; Winding et al., 2016).
Studies have also demonstrated that actin filaments contribute to membrane protrusions, working in combination with stable MTs to initiate neurite outgrowth (Dent et al., 2011; Sainath and Gallo, 2015). Together, these results suggest that the local increase in actin dynamics, as well as the mechanical forces produced by MT sliding, all play key roles in stimulating neurite formation.
Axon Elongation and BranchingAfter neurite extension, one of the multiple processes becomes the axon, while the others later develop into dendrites (Menon and Gupton, 2016). Stable MTs are essential in axon specification and actively determine the polarity of developing neurons (Conde and Cáceres, 2009; Hoogenraad and Bradke, 2009). Interestingly, the ratio of stable to dynamic MTs was found to be significantly higher in one specific neurite compared to other neurites during this stage of neural development (Witte et al., 2008). Furthermore, axon formation was induced in unpolarized neurons following the addition of a photoactivatable analog of the MT-stabilizing drug taxol, while the addition of low doses of taxol led to the formation of multiple axons (Witte et al., 2008). These results suggest that in unpolarized neurons, the stabilization of MTs within one specific neurite occurs before axon formation begins. Once neurons begin to establish a distinct polarity, the centrosome progressively loses its function as an MTOC, with MTs beginning to nucleate from non-centrosomal MTOCs, such as Gogli outposts (Stiess et al., 2010; Stiess and Bradke, 2011; Ori-McKenney et al., 2012; Yau et al., 2014). Moreover, the stabilization of non-centrosomal MTs by the minus-end binding protein, calmodulin-regulated spectrin-associated protein 2 (CAMSAP2), was shown to be required for the establishment of neuronal polarity and axon formation (Yau et al., 2014). Reduction of CAMSAP2 inhibited both proper polarization and axon formation in vitro, and led to defects in neural migration in vivo, providing further evidence that MT stabilization is critical for these processes during neural development (Yau et al., 2014). Increased MT stabilization may also provide tracks for selective targeting of MT motor proteins to aid in the transportation of various organelles and proteins that are necessary for the eventual formation of axonal segments (Kapitein and Hoogenraad, 2011). Kinesin-1 has been shown to preferentially bind to stabilized MTs and accumulate in the future axon, contributing to early polarized trafficking, suggesting that an increase of stable MTs may lead to increased kinesin-mediated transport, driving both neuronal polarization and eventual axon specification (Nakata and Hirokawa, 2003; Jacobson et al., 2006).
A major change that occurs, following axon specification, is the formation and enlargement of the growth cone, a dynamic structure at the tip of the growing axon responsible for driving axon elongation and branching (Kahn and Baas, 2016). Neuronal growth cones probe the extracellular environment and come into contact with various external stimuli, thus steering the axon in a particular direction (Lowery and Van Vactor, 2009). The growth cone uses the cytoskeletal machinery to progress through three distinct morphological stages, termed protrusion, engorgement and consolidation (Lowery and Van Vactor, 2009).
To progress through these stages, an array of dynamic MTs penetrate into the central and peripheral domains of the growth cone, and can display different behaviors which include splaying, looping and bundling (Tanaka and Kirschner, 1991). This remodeling is essential for MTs to probe the growth cone periphery in search of guidance cues, which prompt growth cone advancement and turning. Additionally, growth cone formation and advancement require the interaction between actin filaments and dynamic MTs, which work in combination to promote the extension of lamellipodia and filopodia at the tip of the axon (Dent et al., 2011).
Cytoskeletal remodeling must also occur during axon branching, which involves an accumulation of actin filaments that will form axonal filopodia along the axon shaft (Mattila and Lappalainen, 2008). Shortly thereafter, MTs begin to invade the actin-rich filopodia with localized splaying of the normally bundled MT array (Dent et al., 1999; Gallo and Letourneau, 1999). This invasion by the axonal MTs into the filopodia allows their maturation into collateral branches as they continue extending.
Early studies showed that there was an increase in the number of MTs at regions that eventually become axon branches, suggesting that MTs within the parent axon undergo fragmentation, and a portion of these fragments are then translocated to the developing branches (Yu et al., 1994; Kalil and Dent, 2014; Armijo-Weingart and Gallo, 2017). Thus, MT fragmentation and transportation appear to be key mechanisms which regulate the beginning of axon branch formation. Crosstalk between actin and MTs also seems to be required during the initial steps of axon branching (Dent and Kalil, 2001; Kalil and Dent, 2014; Gallo, 2016; Pacheco and Gallo, 2016). Dynamic MTs colocalize with F-actin in regions of axon branching, whereas stable MTs are excluded from these regions (Dent and Kalil, 2001). Moreover, when MT dynamics were dampened in neurons treated with either taxol or nocodazole, invasion of MTs into filopodia was reduced and they were unable to interact with actin-filament bundles, which resulted in decreased neurite formation (Dent et al., 2007).
Recently, the cytoskeleton-associated protein, drebrin, was shown to regulate both actin filaments and MTs to initiate the formation of axon branches (Ketschek et al., 2016; Zhao et al., 2017). Endogenous drebrin was found to localize to axon actin patches in vivo, which eventually form axonal filopodia, suggesting that drebrin may contribute to the development of these precursor structures before axon branching (Ketschek et al., 2016).
Additionally, reduction of drebrin severely inhibited axonal filopodia formation and the number of collateral branches in vitro, further demonstrating an essential role for drebrin in regulating these processes (Ketschek et al., 2016). Expression of drebrin also increased the number of axonal filopodia that contained end-binding protein 3 (EB3) comets, indicating that drebrin can promote the entry of dynamic MTs into axonal filopodia during the formation of axon branches (Ketschek et al., 2016). Thus, dynamic MTs and their interactions with actin filaments are crucial for the establishment and formation of collateral branches during neuronal development.
Following axon formation, other neurites begin to develop into dendrites, prompting dramatic changes to the MT network. While dendrites branch more extensively than axons, the behavior of MTs during this process has not yet been fully elucidated. One of the most striking features that distinguishes dendrites from axons is the orientation of their MTs. Axons display uniform plus-end distal MTs, while dendrites harbor a population of MTs with mixed polarity, where both plus and minus ends are oriented towards the cell body (Kapitein and Hoogenraad, 2015; Tas et al., 2017).
These distinct polarity patterns are essential for determining the directionality of intracellular cargo transport, which maintains both the composition and morphological differences between axons and dendrites. Several studies have suggested that the translocation of MTs of differing orientations within dendrites plays a key role in establishing their mixed polarity (Sharp et al., 1995, 1997; Yu et al., 2000; Zheng et al., 2008; Rao et al., 2017). When MT assembly was inhibited with low levels of vinblastine, dendritic elongation was reduced, however MT reorientation was not hindered, suggesting that transport of MTs from the cell body is a possible mechanism for creating the non-uniform orientation of dendritic MTs (Sharp et al., 1995).
Moreover, several studies have shown that neurons use distinct motor proteins to engage in specific transportation events that may drive this difference in MT orientation between axons and dendrites (Sharp et al., 1997; Yu et al., 2000; Zheng et al., 2008; Rao et al., 2017). Reduction of the kinesin-related motor protein CHO1 (also known as KIF23) from cultured neurons inhibited the movement of minus-end distal MTs into nascent dendrites and these processes failed to differentiate (Sharp et al., 1997; Yu et al., 2000). Likewise, reduction of kinesin-12 (also known as KIF15) produced similar phenotypic results (Lin et al., 2012). The abnormalities observed with depletion of kinesin-6 could be rescued by overexpression of kinesin-12, indicating that these two motor proteins may share functional redundancy within dendrites (Lin et al., 2012).
Similarly, neurons treated with the dynein inhibitor, Ciliobrevin D, displayed a decrease in MT transport and abnormal MT orientation in the axon, which could be rescued after Ciliobrevin D washout (Rao et al., 2017). Thus, active MT transport by specific motor proteins is essential for both establishment and maintenance of MT bi-directionality during dendritic development.
The next steps in neuronal development are the formation and maturation of dendritic spines, which represent postsynaptic sites of excitatory synapses. The connections between an axon and a dendrite of neighboring neurons is a process that continuously occurs throughout development and into adulthood, as the brain rewires these connections in response to novel stimuli. For some time, it was thought that dendritic spines were devoid of dynamic MTs, and that actin was the main regulator of spine morphology and dynamics associated with synaptic plasticity. However, within the last decade, the use of new visualization techniques has revealed that MT dynamics do play an essential role in dendritic spine development (Yau et al., 2016; Dent, 2017).
In concert with MT transport, MT polymerization actively occurs and contributes to the development of the dendritic branches. MT assembly occurs within the cell body, and after transport, these newly formed MTs are incorporated into the dendrite (Dent, 2017). Dynamic MTs can penetrate into dendritic spines of different shapes, including mushroom, stubby and thin, and can modulate their morphology by interacting with F-actin via +TIPs, such as EB3 (Gu et al., 2008; Jaworski et al., 2009; Dent, 2017).
Furthermore, reduction of EB3 impairs spine development, and pharmacological manipulation of MT dynamics severely decreases the total number of spines and inhibits the formation of spines that are induced by brain-derived neurotrophic factor (BDNF; Gu et al., 2008; Jaworski et al., 2009). Interestingly, N-methyl-D-aspartate receptor (NMDAR)-dependent synaptic activation in hippocampal cell cultures increases the proportion of dendritic spines containing dynamic MTs, which contributes to spine enlargement, and this increase in MT invasion is inhibited by either blocking action potential activity or by dampening MT dynamics (Merriam et al., 2011).
Additionally, dendritic spines exhibiting elevations in calcium signaling contain increased amounts of F-actin, and these spines are preferentially targeted by dynamic MTs, which interact with F-actin in a drebrin-dependent manner (Merriam et al., 2013). Moreover, studies have shown that the invasion of dynamic MTs into dendritic spines provide tracks for MT-dependent motors to deliver specific cargoes that are essential for synaptic plasticity (McVicker et al., 2016).
ScopeIt has also been demonstrated that the severing of dynamic MTs by the MT-severing proteins, katanin and fidgetin, are essential for dendrite development and synapse formation (Mao et al., 2014; Leo et al., 2015), which will be discussed in greater detail in subsequent sections. Together, these results strongly indicate that dynamic MTs are key regulators of dendritic spine formation, maintenance and synaptic activity, and that +TIPs play a role in this dynamicity to modulate spine morphology during neural development.
https://www.frontiersin.org/articles/10.3389/fncel.2018.00165/fullFrontiers in Cellular Neuroscience is a leading journal in its field, publishing rigorously peer-reviewed research that advances our understanding of the cellular mechanisms underlying cell function in the nervous system across all species.
Read more.....
Microtubules (MTs) form a rapidly adaptable network of filaments that radiate throughout the cell. These dynamic arrays facilitate a wide range of cellular processes, including the capture, transport, and spatial organization of cargos and organelles, as well as changes in cell shape, polarity, and motility. Nucleating from MT-organizing centers, including but by no means limited to the centrosome, MTs undergo rapid transitions through phases of growth, pause, and catastrophe, continuously exploring and adapting to the intracellular environment. Subsets of MTs can become stabilized in response to environmental cues, acquiring distinguishing posttranslational modifications and performing discrete functions as specialized tracks for cargo trafficking.
The dynamic behavior and organization of the MT array is regulated by MT-associated proteins (MAPs), which include a subset of highly specialized plus-end-tracking proteins (+TIPs) that respond to signaling cues to alter MT behavior. As pathogenic cargos, viruses require MTs to transport to and from their intracellular sites of replication.
https://jvi.asm.org/content/91/16/e00538-17While interactions with and functions for MT motor proteins are well characterized and extensively reviewed for many viruses, this review focuses on MT filaments themselves. Changes in the spatial organization and dynamics of the MT array, mediated by virus- or host-induced changes to MT regulatory proteins, not only play a central role in the intracellular transport of virus particles but also regulate a wider range of processes critical to the outcome of infection.
Yes. They play a role in cell division. So do chromosomes. So do centromeres. So do kinetochores. So does the nuclear membrane. Eliminate any of those and cell division cannot take place. Heck, without the element phosphorous cell division could not take place.We disagree. Microtubules provide the mitotic spindle for all cell division (especially neural cells) in the first place .
No you are not recognizing that all the complexity of all cells are based on the cytoskeleton network which consist of three chemical structures, the microtubules , microfilaments, and intermediate filaments. The "cytoskeleton" is responsible for the growth, shape, coherence, and function of all those individual cells and organelles.That title is not held by microtubules. It is held by dozens of elements, compounds and structures within the cell. Your obsession with microtubules has blinded you to the complexity of the process.
What would happen if someone snuck in during the night and stole your skeleton? Just to be clear, that’s not very likely to happen, biologically speaking. But if it did somehow happen, the loss of your skeleton would cause your body to lose much of its structure. Your external shape would change, some of your internal organs might start moving out of place, and you would probably find it very difficult to walk, talk, or move.
Interestingly enough, the same is true for a cell. We often think about cells as soft, unstructured blobs. But in reality, they are highly structured in much the same way as our own bodies. They have a network of filaments known as the cytoskeleton (literally, “cell skeleton”), which not only supports the plasma membrane and gives the cell an overall shape, but also aids in the correct positioning of organelles, provides tracks for the transport of vesicles, and (in many cell types) allows the cell to move.
What 3 cell parts are made of microtubules?In eukaryotes, there are three types of protein fibers in the cytoskeleton: microfilaments, intermediate filaments, and microtubules. Here, we'll examine each type of filament, as well as some specialized structures related to the cytoskeleton......more
https://www.khanacademy.org/science...-a-cell/tour-of-organelles/a/the-cytoskeletonMicrotubules are also key components of three more specialized eukaryotic cell structures: flagella, cilia and centrosomes.
www.ruf.rice.eduMicrotubules form a framework for structures such as the spindle apparatus that appears during cell division, or the whiplike organelles known as cilia and flagella. Cilia and flagella are the most well-studied models for microtubule structure and assembly, and are often used by textbooks to introduce microtubules.
The fundamental driving structure behind every bit of complexity in the cell is the DNA of the cell, contained in the chromosomes, which is in turn contained in the nucleus. Everything the cell does, every protein it encodes, every structure it creates, comes from the instructions encoded in the cell's DNA. They create the endoplasmic reticulum, the ribosomes - and yes, even the microtubules.No you are not recognizing that all the complexity is gased on the cytoskeleton which is the microtubule network.
Yes, the DNA is the blueprint of the cell shape and function. But Microtubules are responsible for cell division (mitotic spindle) and the copying of the DNA itself.The fundamental driving structure behind every bit of complexity in the cell is the DNA of the cell, contained in the chromosomes, which is in turn contained in the nucleus. Everything the cell does, every protein it encodes, every structure it creates, comes from the instructions encoded in the cell's DNA. They create the endoplasmic reticulum, the ribosomes - and yes, even the microtubules.
Despite differences between prokaryotes and eukaryotes, there are several common features in their cell division processes. Replication of the DNA must occur. Segregation of the "original" and its "replica" follow. Cytokinesis ends the cell division process. Whether the cell was eukaryotic or prokaryotic, these basic events must occur.
Cytokinesis is the process where one cell splits off from its sister cell. It usually occurs after cell division. The Cell Cycle is the sequence of growth, DNA replication, growth and cell division that all cells go through. Beginning after cytokinesis, the daughter cells are quite small and low on ATP. They acquire ATP and increase in size during the G1 phase of Interphase. Most cells are observed in Interphase, the longest part of the cell cycle.
After acquiring sufficient size and ATP, the cells then undergo DNA Synthesis (replication of the original DNA molecules, making identical copies, one "new molecule" eventually destined for each new cell) which occurs during the S phase. Since the formation of new DNA is an energy draining process, the cell undergoes a second growth and energy acquisition stage, the G2 phase. The energy acquired during G2 is used in cell division (in this case mitosis).

Prokaryotes are much simpler in their organization than are eukaryotes. There are a great many more organelles in eukaryotes, also more chromosomes. The usual method of prokaryote cell division is termed binary fission. The prokaryotic chromosome is a single DNA molecule that first replicates, then attaches each copy to a different part of the cell membrane. When the cell begins to pull apart, the replicate and original chromosomes are separated. Following cell splitting (cytokinesis), there are then two cells of identical genetic composition (except for the rare chance of a spontaneous mutation).

https://www2.estrellamountain.edu/faculty/farabee/biobk/BioBookmito.htmlThe prokaryote chromosome is much easier to manipulate than the eukaryotic one. We thus know much more about the location of genes and their control in prokaryotes.
Mitosis is the process of forming (generally) identical daughter cells by replicating and dividing the original chromosomes, in effect making a cellular xerox. Commonly the two processes of cell division are confused. Mitosis deals only with the segregation of the chromosomes and organelles into daughter cells.
Eukaryotic chromosomes occur in the cell in greater numbers than prokaryotic chromosomes. The condensed replicated chromosomes have several points of interest. The kinetochore is the point where microtubules of the spindle apparatus attach. Replicated chromosomes consist of two molecules of DNA (along with their associated histone proteins) known as chromatids. The area where both chromatids are in contact with each other is known as the centromere the kinetochores are on the outer sides of the centromere. Remember that chromosomes are condensed chromatin (DNA plus histone proteins).

During mitosis replicated chromosomes are positioned near the middle of the cytoplasm and then segregated so that each daughter cell receives a copy of the original DNA (if you start with 46 in the parent cell, you should end up with 46 chromosomes in each daughter cell). To do this cells utilize microtubules (referred to as the spindle apparatus) to "pull" chromosomes into each "cell". The microtubules have the 9+2 arrangement discussed earlier. Animal cells (except for a group of worms known as nematodes) have a centriole.
Plants and most other eukaryotic organisms lack centrioles. Prokaryotes, of course, lack spindles and centrioles; the cell membrane assumes this function when it pulls the by-then replicated chromosomes apart during binary fission.
Cells that contain centrioles also have a series of smaller microtubules, the aster, that extend from the centrioles to the cell membrane. The aster is thought to serve as a brace for the functioning of the spindle fibers.

https://www2.estrellamountain.edu/faculty/farabee/biobk/BioBookmito.htmlThe phases of mitosis are sometimes difficult to separate. Remember that the process is a dynamic one, not the static process displayed of necessity in a textbook.......more
Nope. Put a bunch of microtubules in a petri dish and add a bunch of denatured DNA and . . . exactly nothing will happen. No copying. No new proteins. No functioning copy machine.Yes, the DNA is the blueprint of the cell shape and function. But Microtubules are responsible for cell division (mitotic spindle) and the copying of the DNA itself.
Microtubules function as copy machines.
No one is disputing that. Why do you keep repeating something which is not in question.The fundamental driving structure behind every bit of complexity in the cell is the DNA of the cell, contained in the chromosomes, which is in turn contained in the nucleus. Everything the cell does, every protein it encodes, every structure it creates, comes from the instructions encoded in the cell's DNA.
No DNA doesn't do a thing except make copies of itself. It's is a long polimer consisting of four chemicals, which can only duplicate itself. How that works seems to be still unknown.They create the endoplasmic reticulum, the ribosomes - and yes, even the microtubules.
https://www.khanacademy.org/science...dna/a/hs-dna-structure-and-replication-reviewDNA replication is not the same as cell division. Replication occurs before cell division, during the S phase of the cell cycle. However, replication only concerns the production of new DNA strands, not of new cells.


Cytokinesis (/ˌsaɪtoʊkɪˈniːsɪs/) is the part of the cell division process during which the cytoplasm of a single eukaryotic cell divides into two daughter cells. Cytoplasmic division begins during or after the late stages of nuclear division in mitosis and meiosis.
https://en.wikipedia.org/wiki/CytokinesisDuring cytokinesis the spindle apparatus partitions and transports duplicated chromatids into the cytoplasm of the separating daughter cells. It thereby ensures that chromosome number and complement are maintained from one generation to the next and that, except in special cases, the daughter cells will be functional copies of the parent cell. After the completion of the telophase and cytokinesis, each daughter cell enters the interphase of the cell cycle.
DNA does not make copies of itself.No DNA doesn't do a thing except make copies of itself.
DNA is not a polymer consisting of four chemicals. Nor can it duplicate itself. It needs a lot of help.It's is a long polimer consisting of four chemicals, which can only duplicate itself.
Nope. Read the very thing you posted: "The primary enzyme involved in [duplication] is DNA polymerase which joins nucleotides to synthesize the new complementary strand." Nothing about microtubules.It is the microtubules which are the machinery responsible for "cell division".
Now that could be easily tested. And would clear up many questions about the role and function of this little self-assembling dynamical machine.Nope. Put a bunch of microtubules in a petri dish and add a bunch of denatured DNA and . . . exactly nothing will happen. No copying. No new proteins. No functioning copy machine.

https://en.wikipedia.org/wiki/Miller–Urey_experimentAfter Miller's death in 2007, scientists examining sealed vials preserved from the original experiments were able to show that there were actually well over 20 different amino acids produced in Miller's original experiments. That is considerably more than what Miller originally reported, and more than the 20 that naturally occur in the genetic code.[7] More recent evidence suggests that Earth's original atmosphere might have had a composition different from the gas used in the Miller experiment, but prebiotic experiments continue to produce racemic mixtures of simple-to-complex compounds under varying conditions.[8]